第4章 栄養 5.高齢者の時間栄養学
公開月:2020年5月
早稲田大学 先進理工学部電気・情報生命工学科 教授
柴田 重信
1:はじめに
体内時計システムは脳の視交叉上核(SCN)に存在する主時計、大脳皮質や海馬などに存在する脳時計、さらに肝臓、腎臓、副腎、骨格筋などに存在する末梢時計と、3大別できるが、場合によっては主時計と、脳時計と末梢時計を合わせた末梢時計に2大別する。体内時計の性質について述べ、その後それぞれの部位または組織の体内時計に対する加齢の影響を考える。体内時計の研究は時間生物学として発展してきたが、この学問の応用として、薬物の摂取との関係で、「時間薬理学」が、食や栄養の摂取との関係で「時間栄養学」が生まれてきた。ここでは、体内時計と加齢の関係、高齢者と時間栄養のかかわりについて記述する。
2:1日のリズム
1.中枢時計と末梢時計
最も代表的な周期として、約1日を1周期とするサーカディアンリズム(概日リズム)があり、活動量、体温、睡眠・覚醒、摂食など多くの生理機能に関与している(図1)。特に哺乳類においては、代謝、消化・吸収、エネルギー消費を含む多くの生理機能が体内時計によって制御されている1、2)。概日リズムは、細胞内の時計遺伝子(Clock、Bmal1、Cry、Per)の転写・翻訳のフィードバックループにより、mRNAやタンパク質の転写や発現量に日内変動を作り出すと考えられている1、2)。また、体内時計を制御する時計遺伝子(Per1、Clockなど)の発現リズムは、主時計と呼ばれる視交叉上核(SCN)に強力に発現し、脳時計と呼ばれる大脳皮質、海馬、線条体などに、さらには末梢時計と呼ばれる肝臓、膵臓、脂肪、骨格筋などの末梢組織にも強く発現することがわかっている(図1)。これら主時計、脳時計、末梢時計の調和が時計システムとして働くために必要である。体内時計は体温リズムのようにコサイン波で外挿できるようなリズム性を示す現象が多く、周期、振幅、位相を算出することができる。
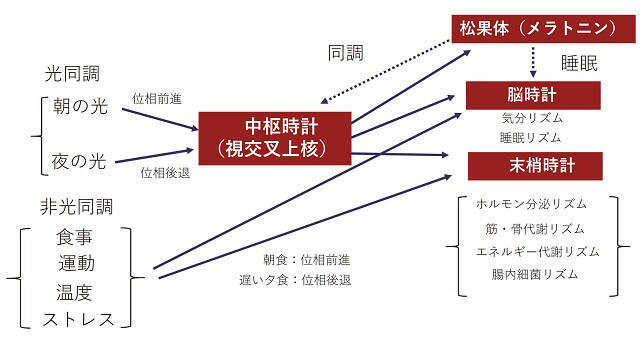
2.同調作用
外界の刺激で24時間に合わせる仕組みがない場合はサーカディアンリズムの周期は種によって、あるいは系統によっても異なるが、おおむね24時間に近い値を示す。このような状態をフリーランと呼ぶ。ヒトの場合、24時間から15-30分程度の長周期を示すことが知られている。明暗状態から、薄暗い明りにしてフリーラン状態で、光パルスを与えると、もともと暗期の中頃だったところ(遅い夕方)に光が当たると、位相が後退し、暗期の終わりだったところ(早朝)に光が当たると位相が前進する。したがってヒトでは、早朝の光で毎日15分から30分、位相を前進させ、地球の24時間周期に合わせている。主時計が目からの光で同調し、その情報が神経や内分泌ホルモンを通して他の脳や末梢臓器の時計遺伝子発現を調節していると考えられた。ところが、毎日一定時刻に餌をあげるとマウスの肝臓や腎臓などの末梢臓器の位相も餌を与えた時刻に依存した位相のリズムを刻む。他の環境要因では、温度変化、運動負荷、ストレスなども同調因子として働く3)。
3:主時計(SCN)と加齢
SCNの時計としての性質や機能が加齢によって変化すると、全身の時間秩序は大きく影響を受けることになる(図2)。時計遺伝子発現リズムの日内変動を調べた研究によると、加齢したマウスやラットでも、基本的には若齢動物と同様に正常なリズムを刻むことが知られている。Per1やPer2などの遺伝子発現を生物発光リズムとして、ex-vivoで評価する方法によると、リズムの振幅は正常か低下傾向にある4)。Per2、Clock、Bmal1の遺伝子発現レベルも低下する5、6)。また、位相に関しては、すこし遅れているという論文や正常という論文があり、周期に関しては少し短縮しているという6)。SCNは光による同調や位相変化が顕著に現れることから、明暗環境を6-8時間前進させたときの新規な位相への反応の違いを調べると、殆ど変わらないか、あるいは早くなるという報告がある6)。SCNへの光同調の影響をPer1やPer2遺伝子の一過性の発現増大を指標に調べた結果、生後約2年の高齢ラットでは、遺伝子発現が減弱した。恐らく網膜からSCNへ至る情報シグナル伝達が加齢により影響を受けたものと考えられる。SCNの神経活動リズムは昼高く、夜低いというリズムを示すが、加齢でリズム振幅が約50%に低下し、ニューロンネットワークが弱くなり、細胞間の同期性が弱くなる6、7、8)。明暗飼育環境の位相前進に対する行動リズム追従の遅れが加齢動物で知られている。恒常暗下での、リズム周期を調べると、ハムスター、ラット、霊長類では加齢により短くなり、マウスでは長くなる6)。
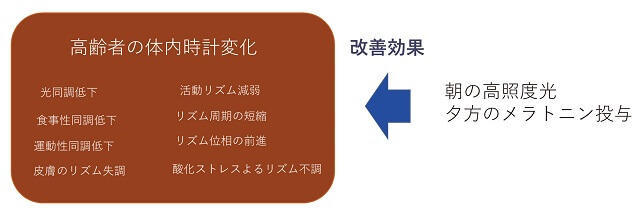
朝の高照度光と、夕方のメラトニンによる改善効果
ヒトの場合、SCN機能を直接調べることはできないので、行動や睡眠覚醒リズムを指標として類推する。ヒトの体内時計周期は若齢者も高齢者も24.18時間だという。ヒトの血圧リズムも振幅が低下し、かつ位相が2時間程度前進する。ヒトの場合、同調させる光の強度が高齢者では若齢者の10倍程度必要だということで、光同調能力が低下している。ヒトの死後脳でSCNの神経を調べた報告では、arginine vasopressin神経が低下していた。また、SCNではないが、死後の大脳皮質のPer1やPer2の時計遺伝子のリズムを、死亡時刻を手がかりに調べた結果、高齢になるとPer1遺伝子発現が低下し、Per2遺伝子発現の位相が前進するという。調べた大脳皮質領域は注意、実行、うつに関連する部分なのでこれらの機能のリズム性の失調にかかわる可能性がある9)。また、メラトニン分泌リズムはヒトのSCNリズムをより直接的に反映していると考えられるが、メラトニン分泌リズムは低下し、位相が前進する。このメラトニン分泌の低下は高齢者の不眠とも関連する。実際、高齢者の不眠に対して、メラトニン受容体のアゴニストであるラメルテオンを使用することも多い(図2)。
SCNの特徴をまとめてみると、時計遺伝子の分子発振機構は老化の影響を受けにくい、一方で、SCNの出力である行動リズムなどは老化の影響を顕著に受ける。同調性の入力機構、発振機構、出力機構のいずれも老化で低下する可能性が示唆される。また、明暗環境がしっかりしている状態ではSCNの時計機構はより健全であるが、恒常暗など明暗の手掛かりが弱い状態では、高齢動物ではリズム性を失いやすくなることが分かった。高齢者は、光の同調効果が弱いので、明暗環境が良い状態で生活をし、海外旅行等の時差ボケにはより注意を払う必要があろう。
4:末梢時計と加齢
末梢時計は、先に述べたようにSCN以外の脳組織や末梢の諸臓器を含むので、一括して取り扱えない場合もあるが、研究が進んでいる肝臓の時計などを中心に説明する。末梢臓器の時計遺伝子発現リズムについては、リズムの振幅が少し低下するがリズム形成ができて、若齢に比較して、老齢動物では位相が2時間程度前進している(図2)10)。また、ex-vivoの研究で、明暗環境の末梢時計の位相変化に対する追従は遅く、光同調に時間を要する。SCNの時計と末梢時計は、ある位相関係をもって時を刻むのが重要であるが、明暗環境シフトに対するSCNと末梢時計の反応が、老齢動物では逆方向に行ったりする10)。ex-vivoによる末梢時計の評価は、生体で起こっていることを外挿することによる知見である。そこで、我々は生きた状態で末梢体内時計を評価するためにin vivoで肝臓、腎臓、顎下腺のPer2リズムをモニターする系を確立し、それを老化研究に応用した11)。明暗環境、恒常暗環境いずれにおいても、末梢臓器の時計遺伝子発現リズムは若齢・高齢マウスで類似し、振幅にも差がなかった。明暗環境の8時間の位相前進に対して、主時計や脳時計を直接反映する行動リズムの前進は高齢マウスでは遅れたが、末梢臓器を5-6日目、9-10日目で調べたところ、リズム位相の前進の大きさには若齢と高齢マウスに差はなかった10)。このように、ex-vivoに比較し、in vivoでの評価では、末梢時計は老化の影響を受けにくいようである。恐らく、ex-vivoでは、カルチャー状態に移行させるときの影響が老化動物では強く出現する可能性がある。以上、まとめると、末梢時計の時計遺伝子発振メカニズムや振幅、光同調などはin vivoで観察する限り、老化の影響は受けにくいといえる。
皮膚は老化とともに、時計遺伝子発現リズムも低下してくる。皮膚の時計遺伝子は、皮膚の老化や傷の回復などにかかわることから、皮膚の体内時計は皮膚の健康維持に重要である。また、紫外線照射による皮膚がんの発症などに時間依存性があり、夜間に細胞分裂が盛んなことから、細胞分裂に対する放射線、紫外線、抗がん剤、ラジカル発生物質などの作用は昼に比較し夜に影響が大きいと考えられている。
5:時間栄養学
SCNが指揮者、臓器が楽器で、時計システムは調和のとれたハーモニーを形成している(図1)1、2)。SCNの主時計は朝の光で一時的に周期を短縮し24.5時間を24時間に合わせ(リセットと呼ぶ)、末梢時計は食事や運動などのタイミングで合わせることがわかっている。時間栄養学は2つの方向性から成り立っている。第一に体内時計が食・栄養の働きを調節する方向性である(図3A)1、2)。第二に食・栄養が主時計、脳時計、末梢時計の周期や振幅のみならず、摂取タイミングに応じて位相をリセットする可能性である(図3B)。時計遺伝子が胃・腸で栄養や食品成分の消化・吸収を、肝臓で分解・再構成を、腎臓で排泄を司っていることから、栄養や食品成分は摂取する時間によって作用強度が異なる可能性である。
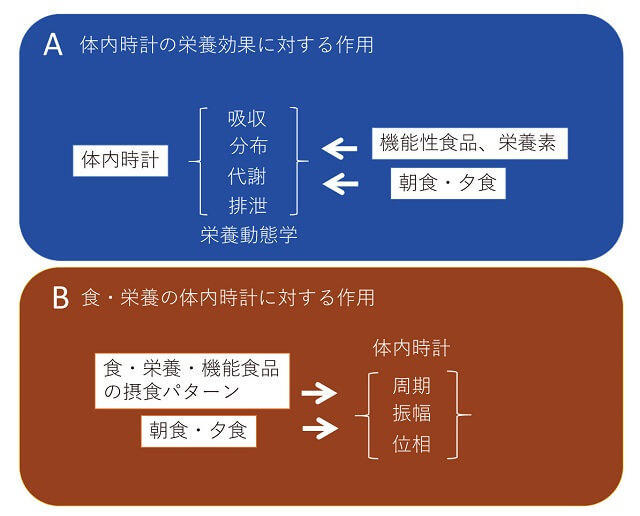
食の体内時計にあたえる影響(A)と、体内時計が食・栄養の効果に与える影響(B)に大別できる
6:「体内時計から食・栄養の働きを調整する方向性」
同一内容の食事を朝、昼、夕、夜中にとった場合の血糖値の推移を調べると、夕食や夜食にとった場合に最大血糖値が高く、元に戻るまでに時間がかかった。したがって、夕食や夜食時の高血糖は脂肪へ変換されやすくなり、肥満のリスクになりやすい。肥満女性の減量プログラムで、1日を1,400kcalとし、朝にウエイトを置いた食事(朝700、昼500、夕200kcal)は、夕にウエイトを置いた食事(200、500、700kcal)に比較して体重や胴回り長が低下する12)。また、朝食に比較して、夕食はエネルギー消費に結びつく食事誘発性熱産生が低いので、遅い夕食や大量の夕食が肥満になりやすい要因の一つになっている。したがって、夕食は低炭水化物が向いているであろう。著者らは、イヌリンやイヌリンが乾燥重量あたり40-50%含まれている菊芋について、朝・夕の摂取タイミングの違いによる影響を24時間モニターの血糖抑制(1週目プレ測定、2週目介入測定)や便通・腸内細菌叢の視点で調べた。朝食時の菊芋5g摂取は夕食時摂取に比較して、24時間血糖値がプレ測定より大きく低下した。さらに、朝食群は、セカンドミール効果で昼食や夕食での血糖上昇も抑制した。一方、夕食の菊芋群では夕食時の血糖は抑制できたが、セカンドミール効果は見られなかった。便通や腸内細菌叢でも、朝食時の菊芋の方が、快便になり、腸内細菌の多様性変化が大きかった。以上、イヌリンやイヌリンを含む菊芋あるいはゴボウなどをデンプン質と一緒に朝食時に取ることは健康維持に寄与する可能性が示唆される。したがって、玄米や精白米に食物繊維を添加した朝食は、1日中の血糖調整や排便に有利に働くことが期待される。
リコピンやDHA・EPAなどの血中濃度は夕摂取より朝摂取で高値が出ることから、これらの機能性食品は朝の摂取が望ましい。また、マウスに高脂肪食や果糖を摂取させると脂肪肝になるが、この時DHA・EPAや魚油を同時に与えると、朝摂取でより効果的であった。DHA・EPAは高齢者の認知機能を改善する効果が知られているので、摂取する場合は、朝がお勧めかもしれない。以上、一般的に、脂溶性が高い物質の経口投与の腸からの吸収には胆汁酸分泌に強く依存する。朝食は胆汁酸分泌が盛んであるので、朝の食事で一緒に脂溶性食品成分を摂取すると、吸収が良いことが分かった。
高齢者のフレイルとノンフレイルの違いが、朝のタンパク質摂取量の違いに起因するという報告がある。アメリカ人、日本人いずれも、また、30代から高齢者まで、全体的に朝食でのタンパク質摂取は少なく、夕食の半分程度である。朝のタンパク質が少ない人は、歩行数が少ないという。高齢者を朝のタンパク質摂取が多い群(朝型)と、夕の摂取量が多い群(夜型)を比較すると、In bodyで調べた筋量や握力は朝摂取群で有意に高値を示した。
7:「食・栄養から体内時計への方向性」
ヒト研究で明暗環境は一定で朝食、昼食、夕食の時間を5時間遅らせると、SCNのメラトニンリズムに変化は見られないが、皮下脂肪の時計遺伝子発現リズムは2.5時間遅れた13)。すなわち主時計と末梢時計が乖離した時差ボケ状態になる。マウスやラットの実験で、長い絶食後の食餌(breakfast)が血糖値を増大させその結果放出されるインスリンシグナルを介して肝臓などの末梢時計をリセットすることが広く知られている1、2)。インスリンはPI3KあるいはMAPKといったリン酸化酵素を活性化させるが、これらの酵素がPER2タンパク質あるいは、BMAL1タンパク質のリン酸化を通して、時計制御にかかわっている可能性が示唆されている。夜の遅い時間の摂食は体内時計の夜型化を助長するが、分食で主食を先に、副食を後に食べることで夜型化を予防できる14)。糖尿病患者のようにインスリン分泌不全や耐性時には、タンパク質食を摂取することにより、IGF-1(インスリン様成長因子)という物質を利用して体内時計をリセットすることができる15)。したがって、デンプン質とタンパク質が含まれるバランスの良い朝食は体内時計のリセットでも有利である。先に朝摂取の菊芋が血糖値上昇を抑制することを述べたが、糖質制限したヒトの糖尿病食でもマウスの肝臓の体内時計は十分にリセットされることから、体内時計のリセットに対してインスリンシグナルは敏感である。したがって、玄米や精白米に食物繊維を添加した朝食でも、末梢体内時計を十分にリセットできる。
高齢者での食事性のリセット効果を調べた研究はないが、高齢マウスでの研究がある。高齢マウスの肝臓、腎臓、顎下腺で、食事性のリセット効果を調べたところ、すべての末梢臓器で、若齢マウスと同定の同調効果を見出すことができた11)。先にも述べたように、高齢者では筋肉維持に朝のタンパク質が重要であることと、またタンパク質を朝食で摂取すれば体内時計のリセット効果も期待でき、高齢者でインスリンの効きが悪い場合も、朝のタンパク質が筋肉維持・同調効果のいずれにも寄与する可能性がある。
文献
プロフィール

- 柴田 重信(しばた しげのぶ)
- 早稲田大学 先進理工学部電気・情報生命工学科 教授
- 最終学歴
- 1976年 九州大学薬学部卒 1981年 九州大学薬学研究科博士課程単位取得退学
- 主な職歴
- 1981年 日本学術振興会奨励研究員 1982年 九州大学薬学部助手、薬学博士(九州大学) 1985年 ニューヨーク州立大学、Research Associate 1995年 九州大学薬学部助教授(薬理学) 1995年 早稲田大学人間科学部助教授 1996年 同・人間科学部教授 2003年 同・理工学部電気・情報生命工学科教授 2006年 同・先進理工学部電気・情報生命工学科教授 2009年 東京農工大学客員教授 2011年 東京女子医科大学大客員教授 現職 早稲田大学先進理工学部電気・情報生命工学科教授
- 専門分野
- マウス、ヒトを研究対象として、体内時計と健康にかかわる分野の研究。特に、薬や食・栄養、運動のタイミングと肥満との関係の研究、あるいは、シフトワークや時差ボケと体内時計の関係やその軽減方法の開発などの研究
- 学会
- 日本時間生物学会理事(副理事長)、時間栄養科学研究会会長
- 賞罰
- 1994年 日本薬学会学術奨励賞受賞
2016年 食創会「第20回安藤百福賞 優秀賞」(公財)安藤スポーツ・食文化振興財団 - 著書
- 2017年 時間栄養学入門、食べる時間を変えれば健康になる ディスカヴァー21 2017年 体内時計健康法 杏林書院
※筆者の所属・役職は執筆当時のもの
PDFダウンロード